interaction of PR70 and the
enhanced binding of PR70 and the
GST-A (Fig. 6A, lanes 3 and 9). This decrease in associated
C-subunit was not observed with the
decrease in C-subunit in the presence of calcium is due to dis-
placement of endogenous regulatory and catalytic subunits
from GST-A by excess free PR70.
recruit additional A- and C-subunits in the presence of calcium.
In contrast, the
units was pulled down with GST-Cdc6 from lysates expressing
the
lysates from non-transfected cells (not shown) suggesting that
GST-Cdc6 can interact with endogenous A- and C-subunits in
the absence of expressed PR70 (presumably by binding to
endogenous PR70). The amounts of A- and C-subunits bound
to GST-Cdc6 in experiments with the
additional support for the conclusion that calcium can regulate
the association of PP2A with Cdc6 and shows that a C-terminal
region of PR70, which includes the PR70-unique domain, is
necessary for interaction with Cdc6.
the phosphatase that dephosphorylates these sites should pro-
mote accumulation of Cdc6. Therefore, RNA interference was
used to determine if knockdown of PR70 affected the levels of
Cdc6. Knocking down the catalytic subunit of PP2A increased
the levels of endogenous Cdc6 in HeLa cells (Fig. 7A). Treat-
ment of cells with either a control siRNA or an siRNA that
knocks down protein phosphatase 5 had no effect on Cdc6 lev-
els. Knocking down the PR70 subunit also caused a substantial
increase in the levels of Cdc6 (Fig. 7B). The increase in Cdc6
levels occurred with two PR70 siRNAs targeted to distinct
regions of the mRNA. The accumulation of Cdc6 following
knockdown with PR70-1 siRNA appeared to be greater than
that with PR70-2 siRNA, which is consistent with the greater
efficiency of the PR70-1 siRNA in reducing PR70 levels (supple-
mental Fig. S2
tion of Cdc6 caused by knockdown of PR70. Knockdown of
PR70 caused an increase in the levels of expressed wild-type
GFP-Cdc6 compared with transfection with a control siRNA
(Fig. 7B). Transfection with a mutant of Cdc6 in which all three
N-terminal phosphorylation sites had been mutated to phos-
pho-mimicking aspartic acid residues (DDD-Cdc6) resulted in
substantially higher levels of expression than those observed
with the wild-type protein as previously reported (15). PR70-1
siRNA had little or no effect on the levels of DDD-Cdc6. The
ability of PR70 knockdown to cause accumulation of Cdc6 was
also greatly diminished when the phosphorylation sites were
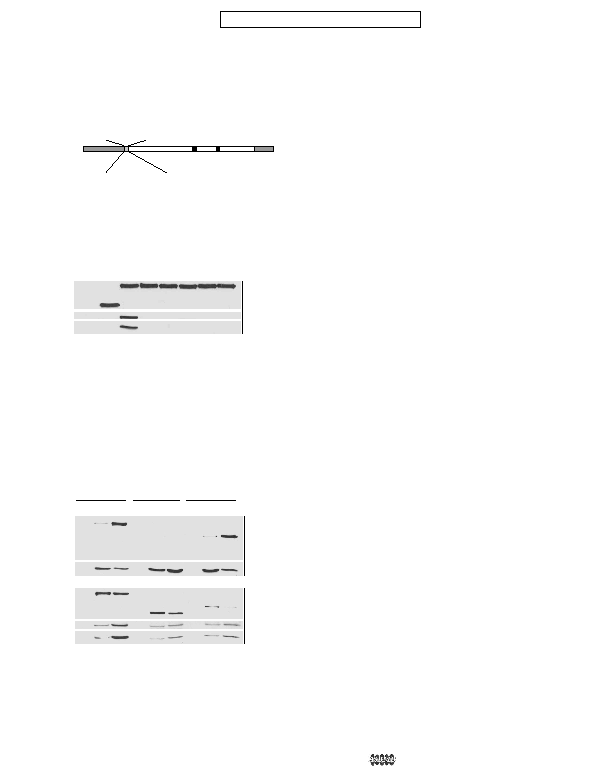
Human PR72
FAF
FYA
AYA
IP
tion of PR70 with PP2A. A, an alignment of the N-terminal region of the
conserved R3 domains (residues 125–135 of PR70) of PPP2R3 subunits
from various species. Conserved FYF residues are shown in bold. B, a dia-
gram showing the residues within the PR70 that were changed in the
mutant forms of PR70 listed on the left. Amino acid substitutions are
shown in bold. C, FLAG-tagged PR70 (PR70), the
cells were lysed, and tagged proteins were immunoprecipitated with anti-
FLAG antibody (Anti-FLAG IP). The immunoprecipitates were resolved by
SDS-PAGE and immunoblotted with anti-FLAG (FLAG), anti-A-subunit (A),
and anti-C-subunit (C) antibodies. A control immunoprecipitate using
lysate from cells transfected with the empty expression vector (Emp Vec) is
shown in the first lane.
pulldown
pulldown
Cdc6. FLAG-PR70 (lanes 1–3), the
were lysed in the presence of EGTA (E) or CaCl
and bound proteins were detected by immunoblotting with anti-FLAG
(FLAG), anti-A-subunit (A) or anti-C-subunit (C) antibodies. Control pull-
downs with GST alone (GST) were carried out with all three expressed
proteins using lysates prepared with standard buffer (lanes 1, 4, and 7).