Cdc6. The potential role of phosphorylation in the effects of
overexpressed PR70 was tested using the phosphorylation site
mutants of Cdc6. Although co-expression of PR70 caused some
increase in the levels of the AAA mutant of Cdc6, the effect was
much less than its effect on wild-type Cdc6 (Fig. 8B). Similarly,
co-expression of PR70 had little effect on the levels of the
DDD mutant of Cdc6 even though endogenous Cdc6 was
increased. Thus, the ability of overexpressed PR70 to cause
accumulation of the protein was inhibited when the phos-
phorylation sites in Cdc6 were mutated. The effects of PR70
overexpression to cause accumulation of Cdc6 suggest it acts
in a dominant-negative manner to block Cdc6 dephospho-
rylation (see “Discussion”).
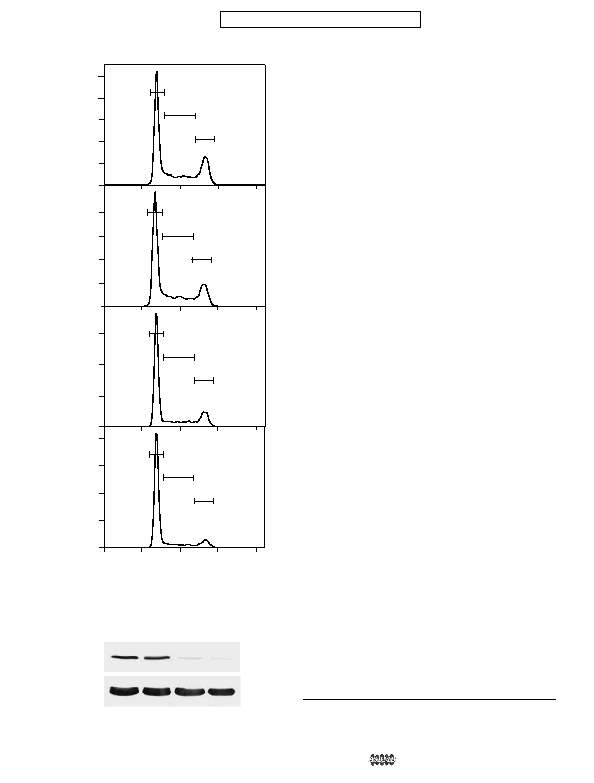
depleted by RNA interference. Knockdown of PR70 caused
accumulation of cells in G
lower levels of PR70 achieved with the PR70 siRNA-1 com-
pared with PR70 siRNA-2 corresponded to a greater increase in
the number of G
subunit in progression through G
of Cdc6. Cyclin-dependent kinases phosphorylate regulatory
sites within the N-terminal domain of Cdc6 and block ubiquiti-
nation by APC/C
of PP2A complexed with the PR70 regulatory subunit as a phys-
iological Cdc6 phosphatase and are consistent with a model in
which PR70 targets PP2A to Cdc6 through direct protein-pro-
tein interactions. Knockdown of PR70 by RNA interference
results in an increase in the levels of Cdc6 protein, consistent
with a role for this subunit in regulating the stability of Cdc6.
Overexpression of PR70 appeared to act in a dominant-nega-
tive manner to also increase the levels of Cdc6. The observa-
tions that increased protein levels did not occur with phospho-
rylation site mutants of Cdc6 are consistent with a role for PR70
in regulating Cdc6 phosphorylation and stability. A novel
aspect of this model is the potential regulation of Cdc6 dephos-
phorylation by calcium. Calcium enhances the recruitment of
the core dimer of PP2A to Cdc6 by binding to the EF-hand
motifs of PR70, raising the possibility that changes in intracel-
lular calcium can regulate the accumulation of Cdc6 and initi-
ation of DNA replication. However, it remains to be deter-
mined if physiological changes in intracellular calcium
PR70-1 siRNA (D). Forty-eight hours later, the cells were harvested and
DNA content determined by FL3 area. The percentages of cells in G