PC7 of the cytoplasmic leaflet), suggesting that 14-carbon acyl
chains are close to the minimum needed to saturate the
hydrophobic belt of AQP0. Furthermore, unlike the lipids in
the DMPC bilayer, several of the longer EPL acyl chains
interdigitate in the middle of the bilayer, filling gaps between
acyl chains in the opposite leaflet (Figure 2B; e.g. one acyl
chain of PE7 inserts between the two acyl chains of PE3 and
the other acyl chain of PE7 inserts between the two acyl
chains of PE4). Other acyl chains bend sharply as they
approach the midpoint of the bilayer and then extend
parallel to the membrane plane (Figure 2B; e.g. acyl chains
of PE4, PE6, and PE7). Such bending may be facilitated by
unsaturated bonds present in acyl chains of EPLs, which have
previously been proposed to explain the bent conformations
seen in lipids associated with cytochrome bc1 complexes
kinks seen in the acyl chains of lipids PE3, PE4, and PE6
occur close to the positions of the double bonds in the most
abundant unsaturated acyl chains of EPLs, 16c1:9 and 18c1:11,
whereas the kink in the acyl chain of PE7 is located between
C6 and C7.
Interaction of the lipid headgroups with AQP0
The AQP0EPL and AQP0DMPC structures make it possible to
analyse the interactions of AQP0 with different lipids.
Although the corresponding lipids in the two bilayers are
at very similar positions (Figure 3C), all the headgroups of
the corresponding lipids in the two bilayers adopt different
conformations. Furthermore, none of the DMPC lipids in the
AQP0DMPC structure interacted with AQP0 through a lipid-
the AQP0EPL structure is in an environment consistent with a
lipid-binding motif: the phosphodiester group of PE3 interacts
with a positivelycharged residue, Arg 196,and a polar residue,
Tyr 105 (Figure 4A). In PC3, the corresponding lipid in the
AQP0DMPC structure (Figure 4B), the glycerol backbone is
shifted by
B3.4 A° closer to the centre of the lipid bilayer.
When Hunte and coworkers identified the lipid-binding motif,
they noted that it did not seem to apply to PC lipids and
suggested that this may be due to the large choline headgroup
were therefore unsure whether the changes in the conforma-
tion of PE3 with respect to PC3 were the result of the presence
of a lipid-binding motif. When we attempted to reposition PC3
to a location more similar to PE3, we found that there was
sufficient space for a choline headgroup to occupy the same
space as the ethanolamine headgroup of PE3. As the choline
headgroup of PC3 does not occupy this position, even though it
would be sterically possible, and therefore does not interact
with AQP0 through Arg 196 and Tyr 105, we conclude that
these two residues do not constitute a true lipid-binding motif.
Together, these observations suggest that the exact chemical
identities of the phospholipid headgroups have a negligible role
in the interaction of annular lipids with membrane proteins,
corroborating previous results obtained by spin-labelling
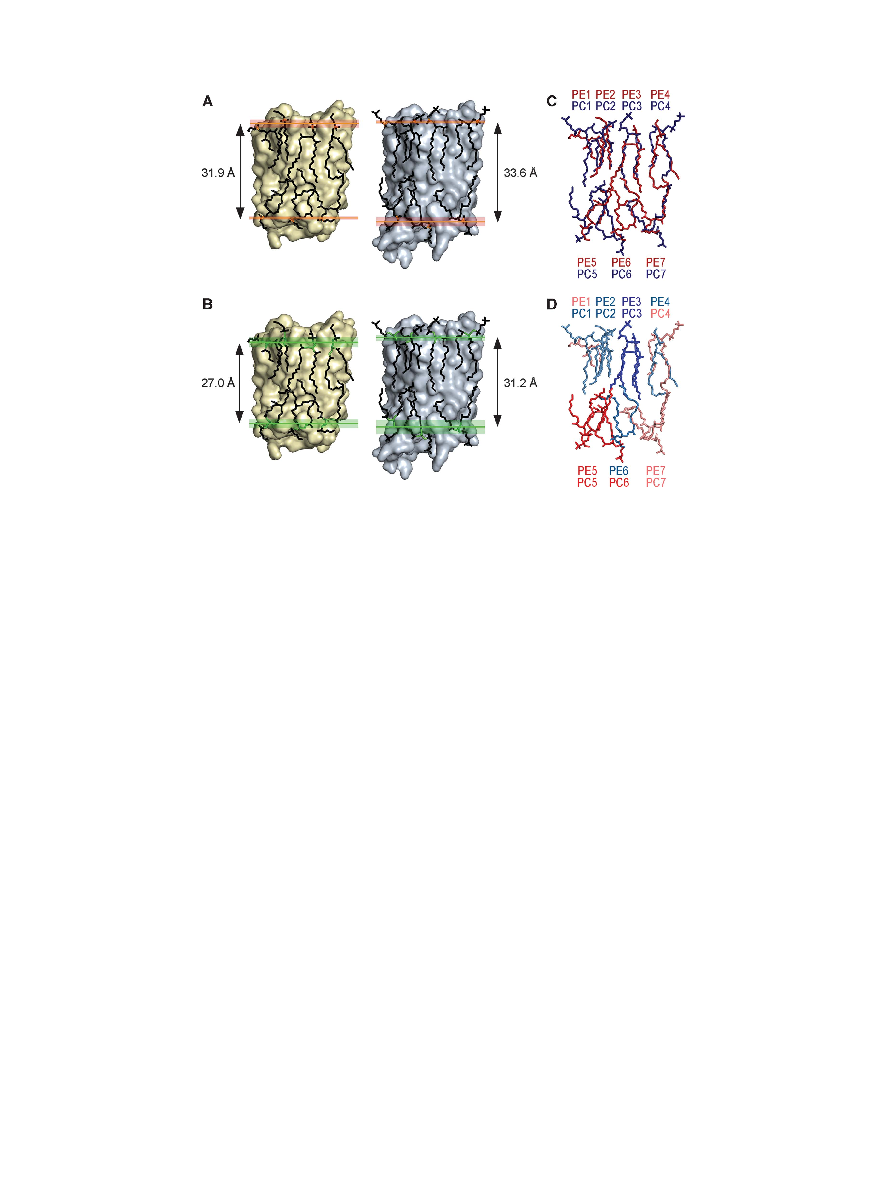
Figure 3 Comparisons between the EPL and DMPC bilayers. (A) The average distance between the phosphodiester groups in the two leaflets of
the bilayer formed by EPLs (31.9 A
° ) is very similar to that between the phosphodiester groups in the DMPC bilayers (33.6 A°). Phosphorous
atoms are shown in orange. Red shading represents the region between the most distantly located phosphorous atoms in each leaflet.
(B) Despite the longer acyl chains of EPLs, the average distance between the C2 atoms of the lipids' glycerol backbones in the two leaflets of the
bilayer formed by EPLs (27.0 A
° ) is shorter than that between the C2 atoms in the DMPC bilayers (31.2 A°). The glycerol groups are shown in
green. The green shading represents the region between the two most distantly located C2 atoms in each leaflet. (C) Overlay of the lipids seen
in the AQP0EPL (red) and AQP0DMPC (blue) structures. (D) Same as in (C) with the lipids coloured according to their B-factors. Colour coding:
intense blue, B-factors below 75 A
° 2; pale blue, B-factors between 75 and 94 A°2; pale red, B-factors between 95 and 114 A°2; intense red, B-factors
above 115 A
° 2.
Interaction of AQPO with E. coli lipids
RK Hite et al
&
2010 European Molecular Biology Organization
The EMBO Journal
VOL 29 | NO 10 | 2010 1655